

En 1717, Nicola Gualtieri describió una extraña concha de un caracol de gran tamaño (puede alcanzar los 55 mm de diámetro de la concha) que había encontrado en España. La concha era excepcionalmente bella y muy diferente de la típica concha de caracoles: era aplanada, fuertemente aquillada y ornamentada con un retículo de costillas espirales y radiales. La descripción llamó la atención de Linneo, que incluyó la especie en su Systema Naturae de 1758 con el nombre Helix gualtieriana, en honor a su descubridor. En 1810, Dennis de Montfort examinó varios ejemplares de esta especie que le habían enviado desde España y consideró que el animal era tan particular que merecía su propio género: el género Iberus, con la especie Iberus gualtierianus. “Nacía” así el que probablemente es el caracol terrestre más singular de los endemismos españoles —de hecho, la revista de la Sociedad Española de Malacología se llama “Iberus”.
En el siglo XIX, los trabajos sobre genitalia de Schmidt (1853) permitieron incluir en el género Iberus otras especies, como Iberus alonensis, las cuales, a pesar de contar con conchas más convencionales, mostraban una genitalia compatible con la de Iberus gualtierianus. De hecho, Iberus gualtierianus e Iberus alonensis pueden hibridar y dar descendencia fértil (García San Nicolás 1957) y tienen cromosomas muy similares (Aparicio 1983), por lo que se llegaron a considerar subespecies de una única especie (de Bartolomé 1982). Pero, ¿cómo pueden ser la misma especie formas conquiológicas tan dispares (véase Figura 1)? Alonso et al. (1985) comprobaron que Iberus gualtierianus se encuentra solo en la Sierra de Gádor (Almería), Sierra Elvira (Granada) y Sierra de Jaén —ahora sabemos que hay otra población en el Valle de Almanzora—, habitando ambientes calizos y xéricos. En base a esto, propusieron que la concha aplanada era un ecotipo que había evolucionado repetidas veces en estas montañas a partir de Iberus alonensis —la especie de Iberus con la distribución más amplia— para permitirles usar las grietas kársticas como refugio durante la estivación y evitar así morir deshidratados.

Con la llegada del siglo XXI se retomó el estudio de la evolución del género Iberus, consiguiéndose respuestas a muchas de las cuestiones planteadas… pero también muchas nuevas preguntas. Mis estudios apoyaron las ideas de Alonso y colaboradores sobre el uso del hábitat por Iberus gualtierianus: el 78% de los ejemplares de esta especie usan las grietas como refugio, los ejemplares que usaron otro tipo de refugios (bajo vegetación o bajo piedras) fueron menos aplanados, y el hábitat de Iberus gualtierianus dentro de Sierra Elvira es más xérico que el de Iberus alonensis (ahora denominado Iberus rhodopeplus), una especie parapátrica de concha globosa con la que llega a hibridar (Moreno-Rueda 2006, 2007, 2011; ver Figura 1).
Los estudios genéticos liderados por Benjamín Gómez-Moliner mostraron que Iberus gualtierianus evolucionó solo una vez, en la Sierra de Gádor, y que su llegada a Sierra Elvira y Sierra de Jaén probablemente fue por vía antrópica (Elejalde et al. 2005). Esto, en una primera lectura, desmontaba la hipótesis de Alonso y colaboradores sobre la evolución iterativa de la morfología de Iberus gualtierianus. Sin embargo, otros resultados apoyaron que, efectivamente, había evolución iterativa de morfologías aplanadas dentro del género Iberus. Aparte de Iberus gualtierianus, otros Iberus presentan concha plana y fuertemente ornamentada: Iberus rositai e Iberus cobosi. Pues bien, los estudios de Elejalde et al. (2008a) demostraron que la forma aplanada ha evolucionado al menos tres veces en el género Iberus.
Además, Elejalde et al. (2008a) encontraron resultados sorprendentes: los caracoles de la especie Iberus cobosi presentan un clado bien definido, pero también había dos “cobosi” mezclados genéticamente con Iberus marmoratus, de concha globosa, lo que se interpretó como un caso de introgresión genética entre ambas especies. Mientras tanto, Iberus rositai no se pudo diferenciar genéticamente de manera clara de Iberus loxanus (de concha globosa), existiendo un gradiente conquiológico entre ambas formas. A estos resultados, más confusos que clarificadores, hay que añadir la tesis de Rodríguez Perochena (2006), que mostró la elevada capacidad de hibridación entre diferentes “especies” del género Iberus, resultado que coincide con las repetidas descripciones de zonas de hibridación entre especies del género.
En los últimos años se han seguido acumulando datos sorprendentes sobre la evolución de este género, especialmente desde el campo de la genética molecular. Próximo al género Iberus existe el género Pseudotachea, que tiene morfología y hábitats bien diferenciados (los Iberus son propios de zonas rocosas, mientras que Pseudotachea habita zonas de matorral). El género incluía dos especies: Pseudotachea splendida y Pseudotachea liturata. Pues bien, los estudios moleculares de Neiber et al. (2021) echaron por tierra esta concepción taxonómica. Resulta que ambas especies pueden integrarse en el clado del género Iberus y, además, en ramas diferentes, lo que sugiere que sus morfologías diferenciales son producto de evolución iterativa en hábitats distintos de los típicos de Iberus, dando lugar —como ocurre con las conchas aplanadas— a evolución convergente.
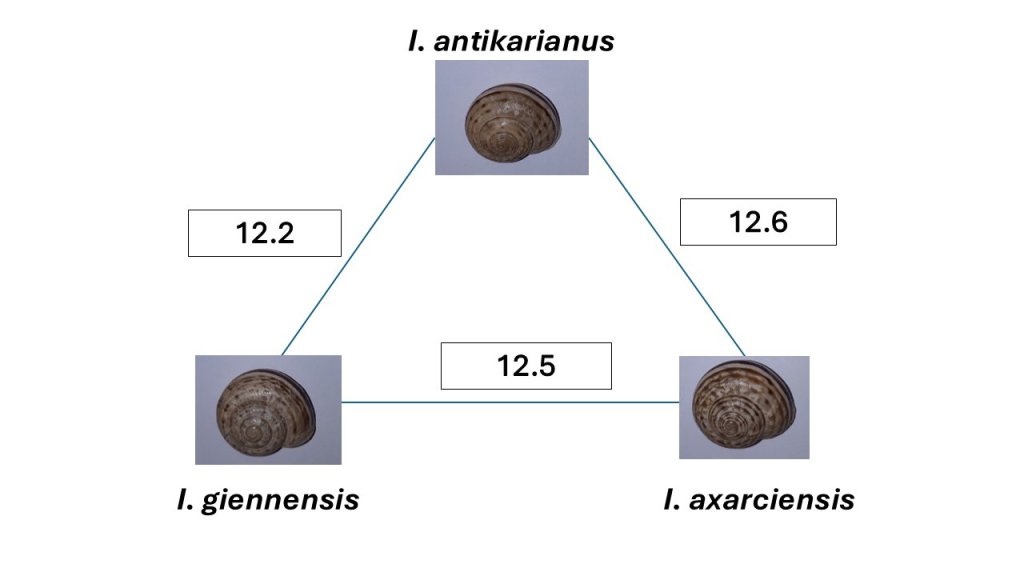
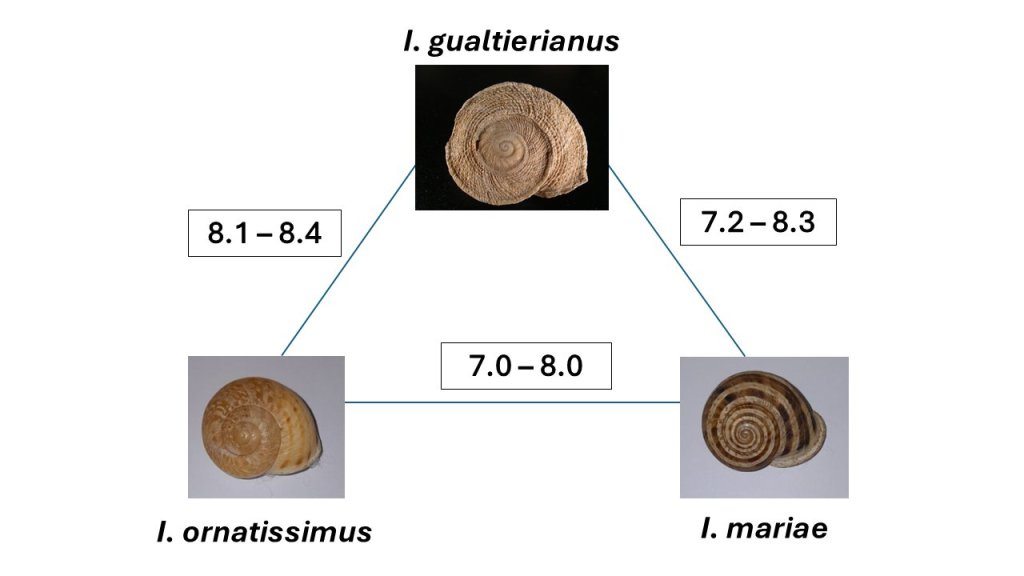
Nuestros recientes estudios muestran, por ejemplo, que existen poblaciones separadas por distancias genéticas superiores al 10%, una distancia genética que justificaría holgadamente considerarlas como especies separadas, pero que apenas se diferencian morfológicamente (Figura 2; Liétor et al. 2024). En cambio, en la Sierra de Gádor coexisten Iberus gualtierianus, Iberus ornatissimus e Iberus mariae, tres “especies” (los taxónomos no terminan de ponerse de acuerdo) con una concha claramente diferenciada (Figura 3), que están separadas por distancias genéticas inferiores al 8.5% (Elejalde et al. 2008b).
Da la sensación de que solo hemos empezado a rascar la superficie de la evolución de este género endémico de la Península Ibérica, y que hemos encontrado más enigmas que respuestas. Aun así, el género Iberus nos ofrece un bello ejemplo de cómo funciona la evolución. En primer lugar, es frecuente la evolución neutral alopátrica (mediada por deriva genética), potenciada por la reducida capacidad de dispersión de los caracoles. Aparentemente, cuando las poblaciones —que suelen habitar en montañas— están aisladas, se produce una diferenciación genética que no se traduce en una importante diferenciación morfológica, probablemente por la reducida implicación de la selección natural en el proceso. Tal es el ejemplo de las especies descritas en Liétor et al. (2024): elevada diversificación genética pero reducida diferenciación conquiológica.
Sin embargo, cuando las poblaciones habitan hábitats diferenciados, la elevada capacidad evolutiva de la concha de estos caracoles se pone de manifiesto. Tal es el caso de Iberus gualtierianus, Iberus mariae e Iberus ornatissimus, muy próximos geográfica y filogenéticamente, pero con una conquiología marcadamente diferente (Elejalde et al. 2008b). En determinadas ocasiones, similares presiones selectivas llevan a una evolución que ocurre de manera repetida para dar lugar a formas similares, como ocurre con Iberus gualtierianus, Iberus rositai e Iberus cobosi (Elejalde et al. 2008a), ecotipos adaptados al aprovechamiento de las grietas kársticas con una evolución independiente; o las especies antes incluidas en Pseudotachea (Neiber et al. 2021), adaptadas a ambientes de matorral. Además, los estudios muestran una elevada capacidad de hibridación, probablemente potenciada por la ausencia de una selección para el aislamiento genético dada la escasa capacidad dispersiva de los caracoles.
En resumen, ¿qué se entrevé en el aparente caos evolutivo de este género? Pues que hay diversos modos de evolución (mediada por la deriva genética o mediada por la selección natural), y que el resultado evolutivo será diferente según el motor evolutivo que intervenga con mayor fuerza. Que las conchas tienen una plasticidad evolutiva muy elevada y, por tanto, una validez taxonómica limitada: no hay correspondencia consistente entre morfología de la concha y relaciones filogenéticas. Que la evolución iterativa ocurre con frecuencia, al menos en este género, dando lugar a formas que convergen cuando enfrentan hábitats similares. Y que hay una elevada hibridación entre “especies”, probablemente por la baja presión selectiva para que evolucione el aislamiento genético. Esto último, además, tiene implicaciones para entender el concepto de especie: un constructo humano que nos permite ordenar epistemológicamente la naturaleza, pero que en la vida real se convierte en un concepto gaseoso, complejo y difícil de definir.
En definitiva, el estudio de la evolución del género Iberus abre varias líneas de investigación inexploradas que nos permitirían entender mejor los detalles y la complejidad del proceso evolutivo.
Autor:
Gregorio Moreno Rueda
Profesor Titular, Departamento de Zoología, Universidad de Granada
Referencias:
1. Alonso M.R., López-Alcántara A., Rivas P., Ibáñez M. (1985). A biogeographic study of Iberus gualtierianus (L.) (Pulmonata: Helicidae). Soosiana 13: 1-10.
2. Aparicio M.T. (1983). The chromosomes of eight species of the subfamily Helicinae (Gastropoda, Pulmonata, Helicidae) from Spain. Malacological Review 16: 71-78.
3. Bartolomé J.F.M. (1982). Comments on some mediterranean rockdwelling helicids. Journal of Conchology 31: 1-6.
4. Elejalde M.A., Muñoz B., Arrébola J.R., Gómez-Moliner B.J. (2005). Phylogenetic relationships of Iberus gualtieranus and I. alonensis (Gastropoda: Helicidae) based on partial mitochondrial 16S rRNA and COI gene sequences. Journal of Molluscan Studies 71: 349-355.
5. Elejalde M.A., Madeira M.J., Arrébola J.R., Muñoz B., Gómez-Moliner B.J. (2008a). Molecular phylogeny, taxonomy and evolution of the land snail genus Iberus (Pulmonata: Helicidae). Journal of Zoological Systematics and Evolutionary Research 46: 193-202.
6. Elejalde M.A., Madeira M.J., Muñoz B., Arrébola J.R., Gómez-Moliner B.J. (2008b). Mitochondrial DNA diversity and taxa delineation in the land snails of the Iberus gualtieranus (Pulmonata, Helicidae) complex. Zoological Journal of the Linnean Society 154: 72-737.
7. García San Nicolás E. (1957). Estudios sobre la biología, la anatomía y la sistemática del género Iberus Montfort, 1810. Boletín de la Real Sociedad Española de Historia Natural 55: 199-390.
8. Gualtieri N. (1717). Index Testacearum Conchyliorum quod ad servantur in Museu Gualtieri exhibentur. Florencia.
9. Liétor J., Tudela A.R., Jódar P.A., Jowers M.J., Moreno-Rueda G. (2024). Slow and steady saves the race: molecular and morphological analysis of three new cryptic species of Iberus land snails from the Iberian Peninsula. Organisms Diversity and Evolution 24: 181-200.
10. Linneo C. (1758). Systema Naturae. 10ª ed. Estocolmo.
11. Monfort D. (1810). Conchyliologie Sistématique et classification méthodique des coquilles, tomo II. París.
12. Moreno-Rueda G. (2006). Selección de hábitat por dos subespecies de Iberus gualtieranus (Gastropoda, Helicidae) en Sierra Elvira (SE de España). Zoologica Baetica 17: 47-58.
13. Moreno-Rueda G. (2007). Refuge selection by two sympatric species of arid-dwelling land snails: Different adaptive strategies to achieve the same objective. Journal of Arid Environments 68: 588-598.
14. Moreno-Rueda G. (2011). Revisión del estatus de conservación del caracol amenazado Iberus gualtieranus gualtieranus. Zoologica Baetica 22: 69-85.
15. Neiber M.T., Chueca L.J., Caro A., Teixeira D., Schlegel K.A., Gómez-Moliner B.J., Walther F., Glaubrecht M., Hausdorf B. (2021). Incorporating palaeogeography into ancestral area estimation can explain the disjunct distribution of land snails in Macaronesia and the Balearic Islands (Helicidae: Allognathini). Molecular Phylogenetics and Evolution 162: 107196.
16. Rodríguez Perochena M.I. (2006). Estudio experimental de la reproducción y el crecimiento de Iberus gualtieranus gualterianus (Linnaeus) e Iberus gualtieranus alonensis (Ferussac) (Gastropoda, Helicidae) en condiciones de laboratorio. Universidad Complutense de Madrid, Madrid.
17. Schmidt A. (1853). Der Geschlechtsapparat der Stylommatophoren in taxonomischer Hinsicht. Berlín.
La fotografía de portada es un ejemplar de Iberus gualtierianus, tomada por Mar Comas en Sierra Elvira (Granada). Las fotografías de la figura 2 son obra del Laboratorio de Colecciones Zoológicas de la UGR (CCZ-UGR). Las fotografías de la figura 3 son obra de Francisco Javier Sánchez Camacho y pertenecen al Centro de Colecciones Científicas de la UAL (CECOUAL).
