

Una de las características más llamativas de las aves, además del vuelo, es su coloración. Las aves muestran una gran variabilidad tanto en el número como en la conspicuidad de los colores que tienen en sus plumajes, algo que ha llamado la atención de la comunidad científica desde que la describieran naturalistas como Darwin, Wallace o von Humboldt.
Originalmente, la existencia de especies con coloraciones vistosas desconcertó a los científicos de la época, ya que estos rasgos parecían no alinearse con los postulados de la teoría de la evolución por selección natural. Tener un plumaje ostentoso y colorido, como la cola de un pavo real Pavo cristatus, no es a priori la mejor manera de pasar desapercibido frente a potenciales depredadores. De hecho, los pavos reales y sus llamativas colas dieron a Darwin algún que otro dolor de cabeza que manifestó en una carta al botánico estadounidense Asa Gray diciendo que en aquellos años la simple visión de un pavo real le ponía enfermo (Darwin 1860).
Nuestro entendimiento sobre la evolución y presencia de estos rasgos comenzó cuando, en 1871, Darwin propuso la teoría de la selección sexual (Darwin 1871). Según esta teoría, la función de rasgos vistosos, como la coloración de muchas aves, no era incrementar la supervivencia de sus poseedores, sino aumentar sus posibilidades de reproducirse, por ejemplo, atrayendo a más o mejores parejas. La selección sexual y sus implicaciones tardaron varios años en ser aceptadas y empleadas por la comunidad científica para explicar la presencia de coloraciones llamativas. Quizás el caso más paradigmático de esta falta de aceptación es el de Abbot H. Thayer, pintor y naturalista norteamericano, quien en 1909 propuso que la función de todas las coloraciones era la de camuflar a sus poseedores (Thayer 1909). Thayer llevó sus ideas al límite, llegando incluso a proponer que el característico color rosado de los flamencos tenía como función el camuflaje (Figura 1).

Hoy en día sabemos que la coloración de las aves cumple toda una serie de funciones que van desde el camuflaje o la termorregulación hasta la señalización de la calidad de sus poseedores en contextos competitivos (Terrill & Shultz 2023). Por lo tanto, la coloración de las aves puede estar modulada tanto por la selección natural como por la selección sexual y, por ende, es esperable que la expresión de rasgos de color sea sensible a los factores ambientales que modulan estas fuerzas evolutivas.
Las especies de aves que encontramos en la naturaleza exhiben una gran diversidad en las coloraciones de su plumaje (Cooney et al. 2022; López-Idiáquez et al. 2025) (Figura 2). Mientras que especies como el herrerillo unicolor Baeolophus inornatus tienen plumajes mayormente homogéneos, otras como el herrerillo común Cyanistes caeruleus o el carbonero sultán Melanochlora sultanea exhiben plumajes mucho más diversos y llamativos. Estudios recientes han mostrado que esta diversidad está en buena medida explicada por las condiciones ambientales que imperan en las áreas de distribución de las especies. Se ha visto, por ejemplo, que especies que habitan en áreas con una mayor productividad primaria (Cooney et al. 2022) y con climas más variables (López-Idiáquez et al. 2025) tienden a mostrar coloraciones más complejas que aquellas que habitan en áreas con una productividad más pobre y climas más estables. Además, también se ha mostrado que el plumaje de las aves no responde como una unidad indivisible, sino que la coloración de las diferentes áreas del plumaje es sensible a diferentes factores ambientales (López-Idiáquez et al. 2025).
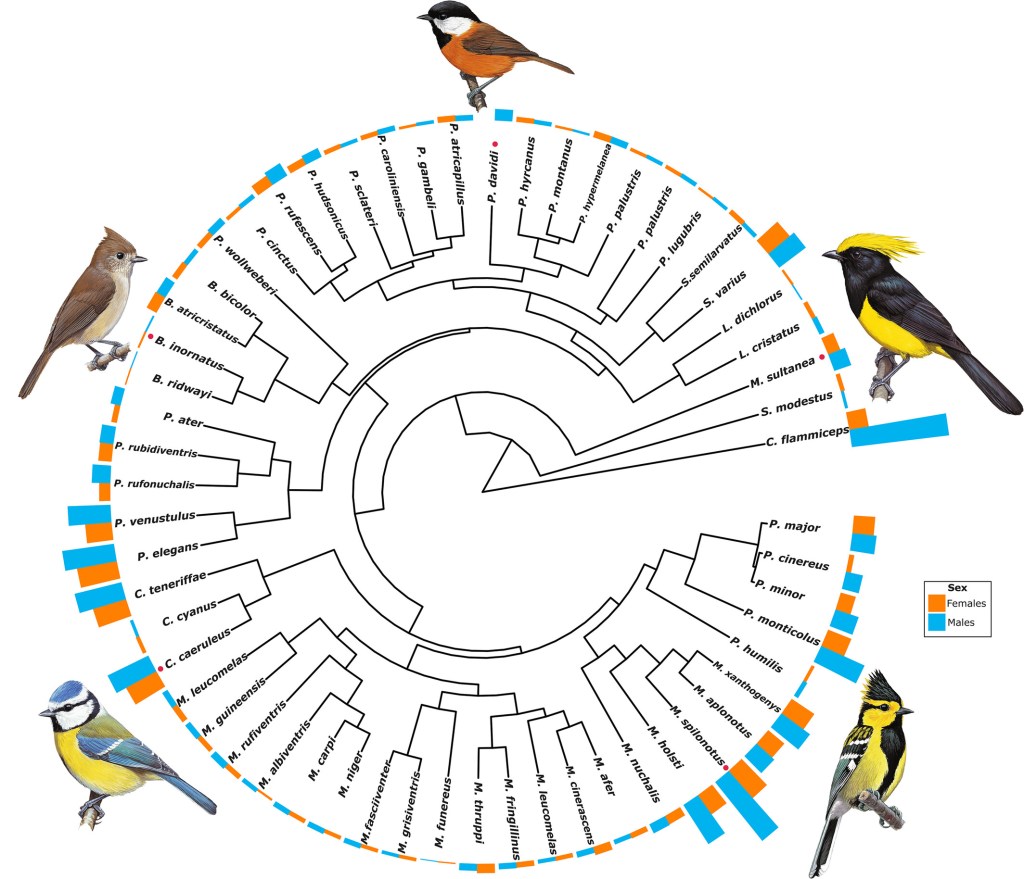
La variabilidad en la coloración no solo ocurre a un nivel interespecífico, sino que también es visible a nivel intraespecífico (López-Idiáquez et al. 2016a, 2016b) (Figura 3). Quizás el caso más paradigmático son las diferencias en coloración entre los machos y hembras de muchas especies de aves (López-Idiáquez et al. 2025). En la mayor parte de casos, estas diferencias radican en el diferente rol que puede tener la coloración en cada sexo. Generalmente, aunque no siempre (Heinsohn 2005), son los machos los que exhiben coloraciones más vistosas que las hembras, ya que las hembras usan los colores de los machos como un indicador de su calidad durante la elección de pareja (Svensson & Wong 2011).
Esta variabilidad intraespecífica también puede estar modulada por la variación en las condiciones ambientales (López-Idiáquez et al. 2016a), ya que la expresión de muchos de los colores que vemos en las aves lleva asociados unos costes de producción o de mantenimiento. Por lo tanto, cuando las condiciones ambientales son buenas, por ejemplo, cuando hay mucha comida, los individuos tienden a mostrar coloraciones más vistosas que cuando las condiciones ambientales son malas (Doutrelant et al. 2012; López-Idiáquez et al. 2016a). Esto se ha visto por ejemplo en las hembras de herrerillo en las que temporadas de cría más duras provocaban que tras mudar exhibieran plumajes más apagados (Doutrelant et al. 2012).
Un caso particular, pero relevante, de efecto ambiental sobre la coloración de las aves es el derivado de las actividades humanas. El cambio climático está generado un impacto sobre la expresión de los colores de aves. Por ejemplo, el incremento de la temperatura asociado al cambio climático ha sido asociado a una reducción en la coloración de los herrerillos (López-Idiáquez et al. 2022) y a una reducción del parche frontal de los papamoscas acollarados (Evans & Gustafsson 2017). Otro ejemplo son las ciudades, el incremento de las superficies urbanas y sus concomitantes cambios ambientales están impactando la coloración de las especies que viven en ellas. Quizás la evidencia más representativa proviene los carboneros comunes Parus major sobre los que existe amplia evidencia que los individuos urbanos presentan coloraciones más apagadas que los individuos que habitan en áreas naturales (Salmón et al. 2023; Sandmeyer et al. 2025).
Mucho se ha avanzado desde que los naturalistas del siglo XIX comenzaron a describir la increíble variabilidad en la coloración de las aves alrededor del mundo. Actualmente sabemos que las condiciones ambientales juegan un papel muy importante a la hora de entender la variación en la coloración entre especies e individuos. No obstante, aún queda mucho por investigar para tener un entendimiento total de cómo se origina esta diversidad y cómo nuestras actividades están generando un impacto sobre uno de los rasgos más llamativos que se puede observar en la naturaleza.
Autor
David López Idiáquez
Investigador posdoctoral en Edward Grey Institute of Field Ornithology, University of Oxford
Referencias:
1. Darwin, C. (1860). Carta de Charles Darwin a Asa Gray el 3 de abril de 1860. Disponible en: https://www.darwinproject.ac.uk/letter?docId=letters/DCP-LETT-2743.xml
2. Darwin, C. (1871). The descent of man and selection in relation to sex. Murray, Londres, Reino Unido.
3. Thayer, A.H. (1909). Concealing-coloration in the animal kingdom. The Mcmillan Co., Estados Unidos.
4. Terrill, R.S. & Shultz, A.J. (2023). Feather function and the evolution of birds. Biological Reviews 98: 540-566.
5. López-Idiáquez, D., Doutrelant, C. & Pearman, P.B. (2025). Global patterns of colouration complexity in the Paridae: effects of climate and species characteristics across body regions. Journal of Animal Ecology 94: 1461-1473.
6. Cooney, C.R., He, Y., Varley, Z.K., Nouri, L.O., Moody, C.J.A., Jardine, M.D., Liker, A., Székely, T. & Thomas, G.H. (2022). Latitudinal gradients in avian colourfulness. Nature Ecology and Evolution 6: 622-629.
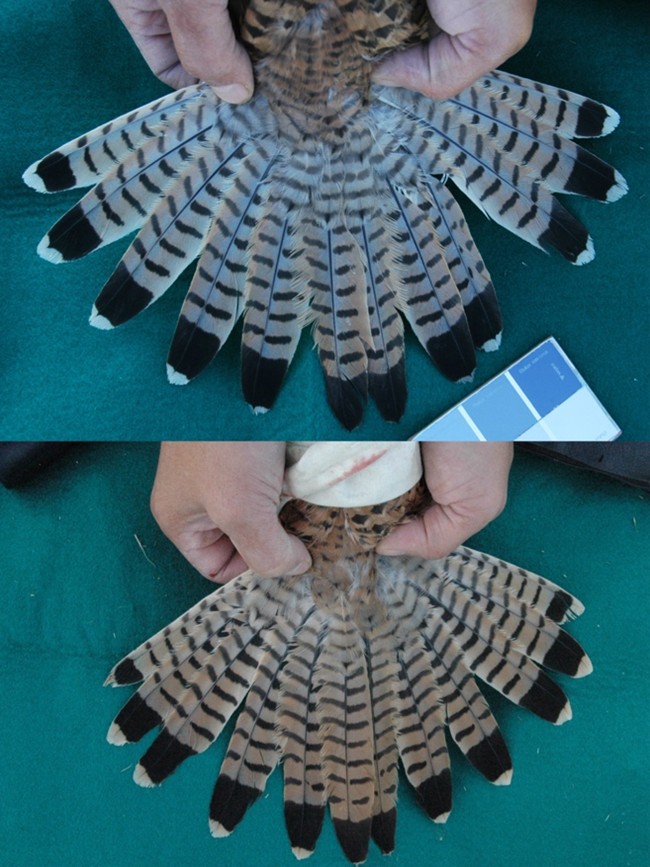
7. López-Idiáquez, D., Vergara, P., Fargallo, J.A. & Martínez-Padilla, J. (2016a). Old males reduce melanin-pigmented traits and increase reproductive outcome under worse environmental conditions in common kestrels. Ecology and Evolution 6: 1224-1235.
8. López-Idiáquez, D., Vergara, P., Fargallo, J.A. & Martínez-Padilla, J. (2016b). Female plumage coloration signals status to conspecifics. Animal Behaviour 121: 101-106.
9. Heinsohn, R. (2005). Extreme reversed sexual dichromatism in a bird without sex role reversal. Science 309: 617-619.
10. Svensson, P.A. & Wong, B.B.M. (2011). Carotenoid-based signals in behavioural ecology: a review. Behaviour 148: 131-189.
11. Doutrelant, C., Grégoire, A., Midamegbe, A., Lambrechts, M.M. & Perret, P. (2012). Female plumage coloration is sensitive to the cost of reproduction. An experiment in blue tits. Journal of Animal Ecology 81: 87-96.
12. López-Idiáquez, D., Teplitsky, C., Grégoire, A., Fargevieille, A., del Rey, M., de Franceschi, C., Charmantier, A. & Doutrelant, C. (2022). Long-term decrease in coloration: a consequence of climate change? American Naturalist 200: 32-47.
13. Evans, S.R. & Gustafsson, L. (2017). Climate change upends selection on ornamentation in a wild bird. Nature Ecology and Evolution 1: 39.
14. Salmón, P., López‐Idiáquez, D., Capilla‐Lasheras, P., Pérez‐Tris, J., Isaksson, C. & Watson, H. (2023). Urbanisation impacts plumage colouration in a songbird across Europe: evidence from a correlational, experimental and meta‐analytical approach. Journal of Animal Ecology 92: 1924-1936.
15. Sandmeyer, L., López-Idiáquez, D., Fargevieille, A., Giovannini, P., Perret, S., del Rey, R., Charmantier, A., Doutrelant, C. & Grégoire, A. (2025). Disentangling urbanisation, climate effects and their interaction on ornamental colourations. bioRxiv.
La imagen de portada, un ocelo de una pluma de macho de pavo real, se ha extraído de Wikimedia Commons (licencia CC 4.0). Los cuadros de los flamencos son obra de A. H. Thayer. La figura del árbol filogenético se ha extraído del artículo López-Idiázquez et al. (2025). Las fotografías de la cola de cernícalo vulgar son obra de Juan Antonio Fargallo.
