

Las ciudades son conocidas por su capacidad para filtrar especies. En concreto, las ciudades favorecen a las especies más adaptadas a los entornos urbanos, induciendo procesos de homogenización biótica. Este proceso de selección actúa a través de los rasgos específicos de las especies (Croci et al. 2008), que están directamente relacionados con las características del paisaje urbano y, en última instancia, determinan las respuestas de las especies a la urbanización.
Sin embargo, el paisaje urbano es heterogéneo. Si atendemos al tipo de desarrollo urbano que se da en las ciudades, este se puede clasificar en dos grupos, que no son más que los extremos de un gradiente (Lin & Fuller 2013). Por un lado se encontrarían las zonas integradoras, que son zonas con baja densidad poblacional, principalmente compuestas por casas unifamiliares y zonas verdes pequeñas y fragmentadas (generalmente en forma de jardines o vegetación en la calle). Por otro lado se encontrarían las zonas separadoras, que poseen una alta densidad poblacional, con bloques de pisos y con extensas zonas verdes bien diferenciadas de las zonas edificadas y no fragmentadas (un ejemplo serían los parques). En estudios anteriores de nuestro grupo de investigación hemos encontrado diferencias en la biodiversidad entre estos dos tipos de desarrollo urbano (Ibáñez-Álamo et al. 2020), pero desconocíamos los mecanismos subyacentes a estas diferencias. Una posibilidad es que estos tipos de desarrollo urbano, dadas sus diferencias en la configuración del paisaje, seleccionen de forma diferente a especies en base a sus rasgos específicos.
Para comprobar esta hipótesis y observar el patrón a escala continental, nuestro grupo de investigación, junto a científicos de otras universidades europeas, tomó datos de las comunidades de aves de 9 ciudades europeas a lo largo del gradiente integrador-separador en cada una de ellas. Realizamos censos en dos estaciones diferentes (primavera e invierno) y extrajimos los datos correspondientes a los rasgos específicos de las especies encontradas en cada ciudad.
Los resultados, publicados recientemente en The Science of the Total Environment, indican que, en efecto, el tipo de desarrollo urbano selecciona unas especies u otras dependiendo de sus rasgos específicos (Ibáñez-Álamo et al. 2024). Además, esta selección ocurre de manera diferente en primavera y en otoño. En primavera, las especies presentes en zonas separadoras se caracterizan por hacer nidos abiertos y tener más inversión reproductora (es decir, tienen mayores tamaños de puesta y número de crías por año), en contraposición a las especies de zonas integradoras, que muestran menor inversión. Estos resultados parecer indicar que las zonas verdes propias de las zonas separadoras, al ser áreas no fragmentadas (normalmente con vegetación nativa), pueden albergar una mayor cantidad y/o calidad de alimento, lo que propiciaría el filtrado de los rasgos de especies menos adaptadas a las perturbaciones humanas. En invierno observamos un patrón de variación similar. En esta estación, las especies presentes en estas zonas separadoras se caracterizaron por exhibir ciclos de vida más cortos en comparación con las especies de las zonas integradoras, donde se encuentran especies más longevas. Estos ciclos de vida longevos, de hecho, también están relacionados con la adaptabilidad de las especies a entornos urbanos.
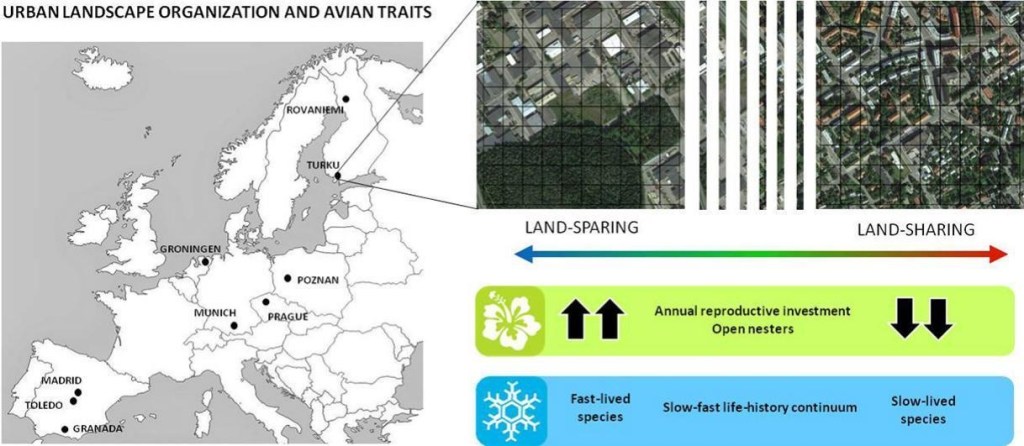
En conjunto, los datos obtenidos en nuestro último estudio indican la necesidad de diseñar ciudades favoreciendo la mezcla e interrelación de ambos tipos de desarrollo urbano, lo que permitiría la existencia de una mayor diversidad de aves. Esto es especialmente importante no sólo desde el punto de vista ecológico (ya que, por ejemplo, se favorecería el funcionamiento de los ecosistemas), sino también para mejorar el bienestar de los ciudadanos.
Autora:
Lucía Izquierdo Extremera
Investigadora predoctoral en el grupo Animal EcoUrban, Universidad de Granada.
Referencias:
1. Croci S., Butet A. & Clergeau P. (2008). Does urbanization filter birds on the basis of their biological traits. Condor 110: 223-240. Doi: 10.1525/COND.2008.8409.
2. Ibáñez-Álamo J.D., Morelli F., Benedetti Y., Rubio E., Jokimäki J., Pérez-Contreras T., Sprau P., Suhonen J., Tryjanowski P., Kaisanlahti-Jokimäki M.L., Møller A.P. & Díaz M. (2020). Biodiversity within the city: effects of land sharing and land sparing urban development on avian diversity. Science of the Total Environment 707: 135477. Doi: 10.1016/j.scitotenv.2019.135477.
3. Ibáñez-Álamo J.D., Izquierdo L., Mourocq E., Benedetti Y., Kaisanlahti-Jokimäki M.L., Jokimäki J., Morelli F., Rubio E., Pérez-Contreras T., Sprau P., Suhonen J., Tryjanowski P. & Díaz M. (2024). Urban landscape organization is associated with species-specific traits in European birds. Science of the Total Environment 908: 167937. Doi: 10.1016/j.scitotenv.2023.167937.
4. Lin B.B. & Fuller R.A. (2013). Sharing or sparing? How should we grow the world’s cities? Journal of Applied Ecology 50: 1161-1168. Doi: 10.1111/1365-2664.12118.
La fotografía que se ha usado como portada (bisbita pratense Anthus pratensis) es obra de Ma Carmen López Luengo y ha sido cedida con permiso de la autora.
